The mammalian cortex displays substantial nonrandom spontaneous neural activity independent of outside sensory input (internally generated).
The question is: How does this internally generated activity influence the formation of new memory representation?
Method related notes:
hM3Dq receptor, is a Gq-coupled receptor which responds to CNO resulting in depolarization and spiking in pyramidal neurons.
Stimulus -----> expression of hM3Dq receptor (in limited to a fraction of excitatory neurons resulting in c-fos promotor drive) -----> So, neurons can be genetically tagged in this way
DOX was used to control in order to capture the receptor expression within a certain time window.
Note: the neural ensemble that got active by this "natural" way, can be reactivated "artificially" again by CNO exposure. This artificially induced activity preserves the spatial character of the neural ensemble. However, it does not preserve the temporal dynamics achieved by natural stimuli (?)
Activity induced by hM3Dq is not temporally coordinated in response to the inducing
stimulus (CNO), as is the case with ChR2-driven
stimulation by light.
Experiment design:

Experiment design:
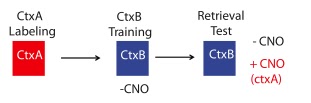
On day 1, mice exposed to CtxA . On day 2, mice got DOX and CNO, then in CtxB got shocked. 24h later tested.
1) Whether the CtxA neurons synthetic activation served as a conditioned stimulus to produce associative fear memory(?)
If so, mice would freeze to CNO or CtxA itself indicating natural and artificial activation of the neurons were similar --> was not the case

Also, CtxA neurons were activated by CNO, and shocked right away in CtxB resulting in no CNO-dependent memory.
Also, when neurons got tagged during conditioning, CNO did not generate freezing.
So, the first option is dismissed.
2) Whether the shocking experience in CtxB would take over and mice would show normal conditioning to CtxB (?)
Mice showed severe freezing deficit to CtxB indicating CNO-induced activity interfere with normal encoding memory for CtxB.
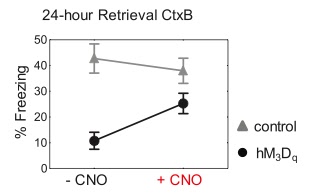
3) A hybrid representation would form as a result of incorporation between CNO-induced artificial stimulation + natural sensory cues from CtxB. (Above) Seeing high freezing to CtxB in CNO+ confirms the third suggestion.
Question: Whether retrieval involves activation of some neurons, which were active during the initial learning (?)
Experiment design:
Result indicates: CNO-induced activation of a competing neural network interferes
with the learned spatial code and degrades recognition if this activity was not present during the
initial training. This is not surprising, given that
even limited focal hippocampal stimulation has
been shown to disrupt spatial memory:

Question: To address ensemble specificity ----> Does the hybrid fear memory formed by
hM3Dqfos mice incorporate the specific pattern of
ctxA neurons activated by CNO during learning,
or are the mice responding to a less specific alteration in brain state?

Result indicates: A requirement for reactivation specifically of the learned (ctxA) neural ensemble, rather than a generalized change in brain state caused by CNO-induced activity.
Supporting design addressing the same above question:

Result : When mice were fear-conditioned after injection of CNO to artificially activate the ctxB ensemble during learning, they developed wild-type levels of 24-hour context fear memory that were independent of CNO stimulation. This is in contrast to the deficit produced in mice pre-exposed to the novel CtxA and further supports the hypothesis that there must be a match in the spatial pattern of neural activity at learning and retrieval.
Paper's conclusions:
- Flexibility in the specific neurons incorporated into a
fear memory trace in the amygdala through a
selection mechanism in which more excitable
neurons are preferentially incorporated into the
trace do not appear to be
attributable to this type of selection, as the reactivation of the neurons with CNO is required
for retrieval, whereas in the previous studies the
stimulated neurons were part of a representation that could be naturally retrieved.
This difference may be due to different requirements for forming simple associations in the amygdala versus more complex representations in the hippocampus and cortex.
This difference may be due to different requirements for forming simple associations in the amygdala versus more complex representations in the hippocampus and cortex.
- They found that the CNO activation alone could not act as an independent cue.
- It is possible that uncoordinated CNO-based stimulus could serve as a conditioned
stimulus if it was limited to a discrete primary sen-
sory area, such as the piriform cortex (?)
- Although the CNO-basedstimulation does not replicate the temporal dynamics of this naturally occurring internal activity,
the approach allows the activation of a distributed spatial pattern of neurons recruited during
a specific experience (ctxA exposure).
No comments:
Post a Comment